2025-11-10
腺相关病毒(AAV)已成基因治疗的关键载体,具备长期表达、安全性与高效递送等优势。本文回溯其从“腺病毒污染物”到临床应用的发展;聚焦分子进化与载体工程(理性设计、定向进化、计算模型),并解析衣壳—基因组相互作用对转导效率与宿主适应的影响。基于343项全球试验,AAV在眼科、神经、肝脏疾病中居核心地位,但仍面临免疫原性、成本与毒性等挑战。此外,AAV库在人类遗体中的应用被报道为理解其在人体内进化与选择的里程碑,为下一代载体优化与临床转化铺路。

AAV:从基础到突破
AAV是一种直径约25纳米的无包膜二十面体病毒,属于依赖辅助病毒(如腺病毒)才能复制的缺陷型细小病毒。其基因组为长约4.7 kb的单链DNA,两端各有一个反向末端重复序列(ITR),作为复制起点并参与基因组包装与持久性。基因组包含REP和CAP两个主要开放阅读框,受p5、p19、p40和p81四个启动子调控。
衣壳由60个蛋白亚基按VP1:VP2:VP3≈1:1:10构成。VP3是主要结构蛋白,可独立形成病毒样颗粒;VP1的N端具有磷脂酶A2结构域,协助病毒逃离内体;VP2则参与衣壳稳定与核转运。这三种蛋白共同决定AAV的组织嗜性,是衣壳工程改造的关键靶点。
AAV高效的基因组结构,结合其独特的复制策略和独特的生物学特性,使其成为人类基因治疗的有前景的候选者。然而直到1982年野生型AAV2型(wtAAV2)感染性克隆被成功克隆到质粒中,这一关键里程碑才得以实现。这一突破标志着传统AAV分子生物学操作的开始,为重组AAV(rAAV)作为基因治疗的传递载体的广泛应用奠定了基础。在接下来的几十年里,rAAV推动了基础研究的重大进展,促进了治疗应用的发展,并促成了诸如Luxturna和Kebilidi等获得美国食品药品监督管理局(FDA)批准的AAV基因治疗产品的开发。
rAAV通过删除REP和CAP基因,替换为治疗用的外源基因,同时保留ITR序列,构建出rAAV载体。这种改造让rAAV彻底失去复制能力,仅保留“递送基因”的功能,安全性大幅提升。如今,我们使用的所有AAV基因治疗药物都源于这一基础设计。
AAV的发展历程:从“污染物”到临床先驱
20世纪60年代
科学家在犬肾细胞(MDCK)中再次发现AAV,确认其“依赖辅助病毒复制”的特性,同时证明它能在不同物种细胞中存活,为后续跨物种应用奠定基础。
20世纪80年代
1980年,Lusby等人确定了AAV2 DNA中ITR的核苷酸序列。他们发现,每个ITR由145个核苷酸组成,且前125个核苷酸通过自配对形成一个T形发夹结构。这种结构特征对于AAV DNA复制至关重要。研究人员还在终端重复区内揭示了核苷酸倒置现象。基于这些发现,他们提出了AAV DNA复制的模型,为之后将其用作基因治疗载体奠定了基础。
1982年,Samulski等人成功克隆了AAV2基因组,从此AAV从一种生物实体转变为人类基因治疗的重要基石。这使对病毒基因组的精确操作成为可能,同时促进了靶向修改和可控载体设计。
1983年,Srivastava等人AAV2提供了首个完整的基因蓝图:该单链DNA基因组长4675个核苷酸,包含145个ITR。研究人员还绘制了三种主要mRNA转录本的5'和3'端,确定了蛋白质合成的潜在起始和终止密码子以及编码三种主要衣壳蛋白的基因片段。
1984年,Hermonat和Muzyczka首次将AAV用于基因传递。研究人员开发了rAAV系统,成功将新霉素抗性基因导入培养的乳腺细胞中。这一开创性的研究证明AAV可用于转导外源DNA,突显了其作为有效基因传递载体的巨大潜力。
20世纪90年代
1993年,Flotte等人研究了rAAV载体在囊性纤维化(CF)基因治疗中的潜力。他们构建了一个携带正常囊性纤维化跨膜传导调节因子(CFTR)cDNA的rAAV载体,并证明了在原代细胞和兔子体内给药后均能成功进行基因转移和表达。在使用AAV-CFTR载体进行治疗后长达6个月时间内,研究人员在受治疗的肺部检测到了载体DNA、CFTR RNA和CFTR蛋白的表达,这为AAV载体能够在体内实现长期稳定的基因表达提供了有力证据。这项研究标志着rAAV在治疗情境下的首次体内应用。
进入21世纪
2001年,McCarty等人开发了自互补型AAV(scAAV)载体:这种载体能够包装二聚体倒置重复基因组,从而绕过了第二链合成的限速步骤。体外和体内实验表明,scAAV载体的转导效率高于传统的单链AAV载体。这些发现突显了scAAV载体在基因治疗中提供更高效转导的巨大潜力,该分子机制已被获批用于脊髓性肌萎缩症(SMA)的药物。
2002年,Rabinowitz等人研究表明,每种AAV血清型都具有独特的组织亲嗜性,这种特性极大地影响了转导效果。通过利用AAV的交叉包装技术,研究人员能够将AAV2的ITR与多种衣壳血清型相结合,从而优化针对特定组织和疾病的基因传递。这种强大的策略提高了靶向特异性,使基于AAV的基因疗法在更广泛的疾病治疗中更具有效性和可及性。此外,交叉包装有助于开发更通用且可扩展的AAV生产系统,简化临床应用的载体制造流程。
2017年,Luxturna成为FDA批准的首款AAV基因治疗药物。该药物用于治疗遗传性视网膜色素变性:通过视网膜下注射rAAV2,递送正常的RPE65基因,让失明患者重见光明。此后,AAV药物进入“爆发期”:2019年Zolgensma(SMA)、2022年Hemgenix(血友病B)、2023年Elevidys(杜氏肌营养不良)、2024年Kebilidi(芳香族L-氨基酸脱羧酶缺乏症)等相继获批,覆盖眼科、神经、血液、肌肉等多个疾病领域。
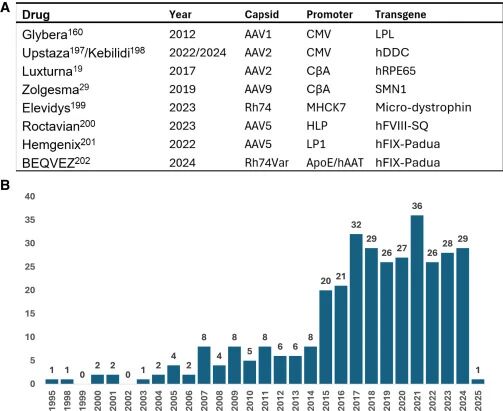
图1:AAV获批药物表与临床试验数量趋势
(图片来源: Lester Suarez-Amaran, et al, Molecular Therapy, 2025)
AAV衣壳设计:从“自然筛选”到“AI 定制”的技术革命
AAV的衣壳是一种高度对称的结构,从宿主细胞的附着到基因组传递的每个病毒生命周期阶段都起着至关重要的作用。它包裹并保护病毒基因组,同时促进多个重要过程,如宿主细胞的进入、细胞内运输、基因释放和免疫逃避等。这些多方面的功能凸显了衣壳在病毒学研究中的重要性,同时也表明其作为一种多功能的治疗性基因传递平台的用途。
在AAV衣壳中,VP3是主要的结构成分。它作为所有三个衣壳蛋白(VP1、VP2和 VP3)共有的骨架,对于维持病毒的整体结构完整性至关重要。值得注意的是,仅表达VP3即足以生成与野生型AAV衣壳结构相似的病毒样颗粒(VLP)。这一能力引发了对替代生产系统的兴趣,如大肠杆菌以及大肠杆菌无细胞蛋白质合成平台等。这些方法旨在降低成本并加快生产速度,使AAV基因疗法更易于获取和大规模应用。
然而,尽管仅使用VP3即可高效生产出衣壳,它们却缺乏由VP1和VP2提供的不可或缺的功能性元素,而这些元素对于病毒的感染性和成功转导至关重要。此外,包括REP蛋白和rAAV基因组在内的AAV包装系统必须适应这些新兴的生产策略。将这些必要的功能性元素整合到以VP3为中心的生产平台中仍是一项重大挑战。要实现这一目标,需要进行精细的工程设计以平衡结构效率、感染性和可扩展性,并确保符合临床级生产标准。
天然的AAV亚型作为通用的病毒载体存在固有的局限性,包括对某些细胞类型的转导效率低下、广泛的组织亲嗜性以及易受中和抗体攻击。为解决这些挑战,已开发出三种主要策略来改造改进AAV衣壳:定向进化、理性设计以及一种将这两种方法相结合的组合式生物工程方法。尽管这三种方法都旨在提高载体性能,但它们在方法论、范围和适用性方面存在显著差异。
1. 定向进化:核心思路是“模拟自然选择”——构建包含数百万种AAV变体的“衣壳库”,在动物模型或人体细胞中筛选出“性能最优”的变体。2003年,Perabo和Müller团队分别开发了首个AAV随机肽库:通过在AAV2衣壳的VR-VIII环(表面暴露区域)插入7个随机氨基酸,构建出包含海量变体的库,最终筛选出能靶向整合素受体的AAV2(I-587),首次证明了“人工进化AAV”的可行性。2016年,Gradinaru团队开发的CREATE平台将AAV9衣壳库与Cre依赖的报告基因结合,成功筛选获得了AAV-PHP.B——这种变体穿越血脑屏障的能力比AAV9提升10倍以上,为中枢神经系统(CNS)疾病治疗提供了新工具。后续升级的M-CREATE平台更是实现了“多组织同时筛选”,相继开发出AAV-PHP.eB(靶向全脑)、AAV-PHP.S(靶向脊髓)等变体,已用于阿尔茨海默病、帕金森病的临床试验。2021年,Tabebordbar团队的DELIVER平台则聚焦肌肉疾病:通过在AAV9衣壳插入RGD基序,筛选出MYOAAV家族,静脉注射后对肌肉的靶向性比AAV9提升100倍,且在非人灵长类动物中效果稳定。目前该疗法已进入杜氏肌营养不良(DMD)的临床研究阶段。
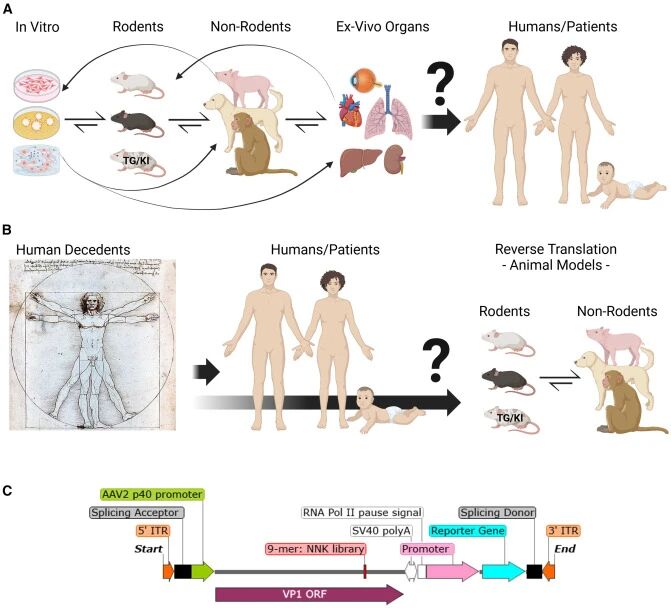
图2:不同AAV进化策略的示意图
(图片来源: Lester Suarez-Amaran, et al, Molecular Therapy, 2025)
2. 理性设计:基于对AAV结构的深入理解,通过定点突变、结构域替换等方式,“按需”优化载体性能。2002年,Xie团队解析了AAV2衣壳的高分辨率晶体结构,首次明确了衣壳表面的受体结合位点,为理性设计提供了蓝图。2008年,Srivastava团队发现AAV2衣壳表面的酪氨酸残基(Y)会被宿主细胞的激酶磷酸化,进而引发泛素化降解,最终降低转导效率。通过将这些酪氨酸突变为苯丙氨酸(F),构建出“Y-F 突变体”(如AAV2(Y444,500,730F)),不仅避免了降解,还将肝脏转导效率提升10倍,且在人体临床试验中安全性良好。2015年,Vandenberghe团队通过“祖先序列重建(ASR)”技术,预测出AAV的“共同祖先”序列Anc80——这种“人造血清型”对肝脏、肌肉、视网膜的靶向性均优于自然血清型,其衍生变体Anc80L65已用于治疗遗传性耳聋的临床试验。
3. AI辅助设计:开启“预测性优化”新时代。随着人工智能技术的发展,AAV载体设计进入“数据驱动”阶段。2019年,Church团队通过对AAV衣壳进行“饱和突变”,绘制出首个AAV衣壳适应性图谱,发现了一个此前未知的基因——MAAP,它能通过“竞争抑制”调控AAV的产量。这一发现为优化载体生产提供了新靶点。2021年,Kelsic团队利用深度神经网络,一次性设计出20万个AAV2衣壳变体,其中5.7万个变体的多样性超过自然血清型,且90%以上能正常组装。更令人惊喜的是,这些AI设计的变体在人类肝细胞中的转导效率比自然AAV2提升50-100倍。2024年,Gradinaru团队开发的APPRAISE平台,通过AlphaFold-Multimer工具预测AAV衣壳与受体的结合结构,再结合生物物理特征评分,能快速筛选出“高亲和力”变体。该平台已成功设计出靶向转铁蛋白受体(TfR1)的AAV变体,静脉注射后能高效穿越血脑屏障,为脑部疾病治疗提供了新工具。同年,Deverman团队的Fit4Function平台则实现了“多性状同步优化”—— 通过机器学习预测AAV的肝脏靶向性、产量、安全性,设计出的变体在非人灵长类动物中,肝细胞转导效率比AAV8提升1000倍,且无明显肝毒性。
AAV设计未来的发展方向与挑战
设计治疗性载体通常需要将不同的组成部分组合在一起,例如启动子、增强子、内含子外显子区域(UTR)、polyA以及其他调控元件。理解调控元件之间的相互作用对于载体工程至关重要。此外,除了调控元件之外,蛋白质编码区域也可以作为转录因子在人类细胞中的结合区域。因此,尽管编码优化已成为一种“常见”的做法,但它也为预测载体性能带来了额外的变数。近来去除CpG二核苷酸的努力往往只是导致另一种形式的密码子优化,而对CpG在AAV生物学中的作用几乎没有影响。事实上,AAV与其他小病毒的区别在于其具有类似CpG岛的基因组结构。野生型AAV具有高GC含量(>50%)以及较高的观察/预期CpG比率(>70%),去除CpG核苷酸片段的做法与野生型AAV中这些元素的进化稳定性相悖,这表明从AAV载体中去除CpG元件时存在一种误导性的做法。无论如何,必须全面考虑所有这些遗传元素对载体性能的综合影响,以尽量减少意外影响。
除遗传元素之间的相互作用外,过去十年的研究还增进了我们对复杂的启动子-衣壳相互作用的理解。在此我们讨论这种相互作用,并强调必须综合考虑所有这些独立的组成部分,以确保载体性能。从历史上看,衣壳(特别是VP3)被认为完全决定了AAV的细胞亲嗜性,导致衣壳工程主要集中在VP3上。然而早期研究提供的证据表明VP1和VP2也会影响细胞特异性表达。传统上认为,当AAV结构蛋白成功将货物送入细胞并完成解旋后,该结构蛋白便完成其功能,而细胞特异性启动子或增强子则会接手并单独决定转基因的表达。然而,此前的研究发现一些结构蛋白突变体能够完成感染的所有步骤;但与野生型结构蛋白相比,其在mRNA积累方面存在显著缺陷。这表明AAV结构蛋白可能在顺式调控基因转录方面发挥作用。
最近,研究者们首次明确了结构蛋白、启动子以及两者之间的相互作用共同决定了细胞特异性转导。例如,在2002年,研究人员使用AAV将在巨细胞病毒立即早期启动子(cytomegalovirus immediate-early promoter,CMV)控制癫痫发作的基因进行传递,展示了显著的启动子相关嗜性以及有效的治疗效果。令人意外的是,在最小的CMV启动子前面添加一个调节元件作为开关却导致了完全相反的结果,使癫痫发作恶化。进一步的分析表明,尽管使用的是相同的衣壳和基因,但假定启动子序列中的一些细微差异(如添加一个小型的转录激活开关元件)导致了抑制性神经元而非兴奋性神经元中转基因的高表达。
临床中AAV载体的安全性、有效性及临床成功率
当前AAV主要的研发重点仍集中在眼部领域,其次是中枢神经系统,肝脏则位居第三,占比分别为26%、21%和18%。这种试验分布表明在过去两年中适应症选择策略并未发生重大变化。这些观察结果表明,当前AAV载体的临床问题(如肝脏毒性)很可能会因器官而异。除非在各个领域完成足够的研究并进行比较以确定其共性,否则这种情况很可能会持续存在。
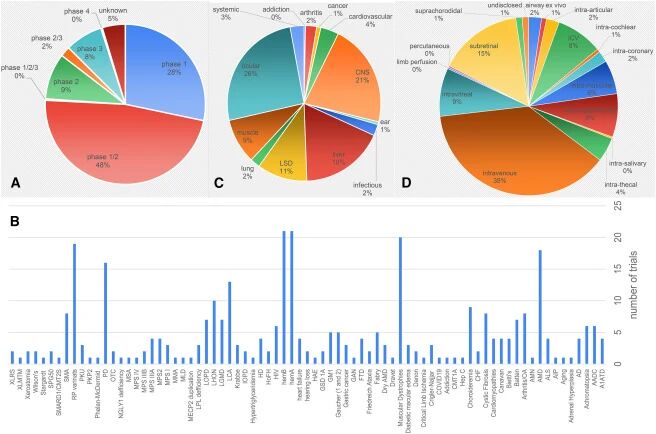
图3:AAV临床试验的描述
(图片来源: Lester Suarez-Amaran, et al, Molecular Therapy, 2025)
目前已有多种给药途径在各种试验中被使用,在此总结在图3D中。与目标组织相一致的是,静脉/血管给药方式仍占主导地位,其次是针对眼部应用的三种给药途径以及用于将药物输送到中枢神经系统的三种途径。根据我们目前对AAV与细胞蛋白质相互作用的理解,给药途径可能会因该环境特有的宿主蛋白质而得到增强或限制。值得注意的是,有四项试验采用了双重给药途径;这些途径未在饼状图中列出。这些双重给药途径可能需要为每种给药途径分别使用不同的衣壳蛋白,从而增加生产成本、毒理学研究等的费用。
本文接下来讨论的最后一个参数是AAV衣壳的利用情况。图4展示了不同衣壳在临床试验中的分布情况。这些试验仍以AAV2衣壳为主,其次是AAV9和AAV8。据Shen等人2022年的报告,当时有20项试验使用了10种独特的工程化衣壳。目前,有39项试验使用了15种独特的工程化衣壳,这表明在过去两年中,新兴的新型衣壳的使用量有了显著增长。
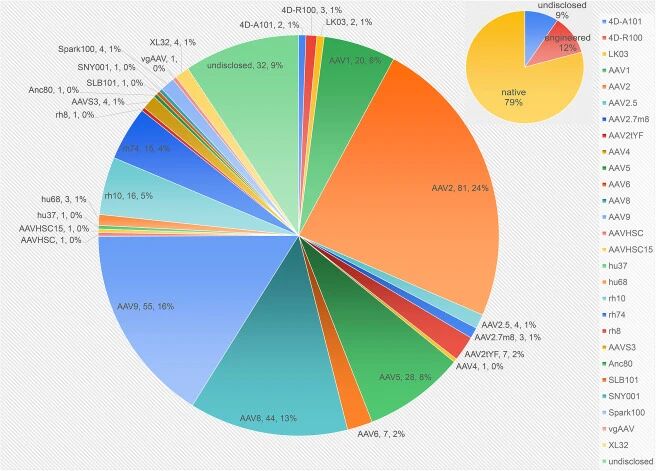
图4:AAV衣壳利用在340个临床试验中的分布
(图片来源: Lester Suarez-Amaran, et al, Molecular Therapy, 2025)
AAV临床试验与经济成效的对比
基因疗法未能如愿广泛应用于单基因疾病。其中一个原因可能是,在动物疾病模型中,我们只关注经过AAV治疗后有所改善的参数,而仅治愈主要症状可能不足以实现最佳的治疗效果。其他原因可能包括对疾病生物学本身的不充分理解、AAV药物生物学、生物标志物缺乏、不合实际需求的试验设计以及对商业预测的错误假设——这对吸引资金来说是一个重要因素。
肝脏导向型基因疗法曾是AAV基因疗法的典范,并被视为典型的“唾手可得的成果”。血友病A和B被视为基因治疗的理想靶点,人们曾预期针对这两种疾病的基因治疗产品将成为重要的畅销药物。用于这些药物的研发投入巨大(总计约数十亿美元),且众多竞争对手开展了多个并行的研发项目(仅列举部分示例:针对血友病B的项目包括辉瑞/Spark、Biomarin、Uniqure/CSL、Freeline Therapeutics、百特/武田、信念医药/武田中国;针对血友病A的项目包括Biomarin、辉瑞/Sangamo、Spark/罗氏、伦敦大学/圣犹达、拜耳/Ultragenix、武田),总计进行了超过40项介入性临床试验以评估。截至文章发表时共有三种获批的血友病药物:针对血友病B的Hemgenix,价格为350万美元;Beqvez/Durveqtix用于血友病B,价格为350万美元;针对血友病A的Roctavian,每位患者的价格为290万美元。到目前为止,Hemgenix和Roctavian的使用情况一直较为缓慢,Beqvez预计也会面临类似的挑战。事实上,Hemgenix的使用速度如此之慢,以至于CSL在其财务报告中并未单独列出其销售额。而Biomarin公司的Roctavian的销售数据分别在2023年(350万美元)和2024年上半年(700万美元)公布,远远低于2023年单一年度1.5亿美元的最初预期。值得注意的是,辉瑞的Beqvez/Durveqtix于2024年中期获得批准,但销售数据尚未公布。讽刺的是,在Beqvez获批后不久,辉瑞决定终止其AAV血友病A项目,尽管其关键试验中的数据表现良好;而罗氏在完成二期研究后也停止了血友病A候选药物的开发。由于前期竞争激烈且获批后使用缓慢,无法确定血友病基因疗法药物是否会成为畅销药,甚至是否能成为主流疗法。与此同时,患者和临床医生对目前获批的药物的接受速度极其缓慢。
而对于美国首次获批的用于治疗LCA的AAV基因疗法——Luxturna(Spark/宾夕法尼亚大学)来说,似乎存在一种中间情况。LCA是一种非致命性疾病,除了已获批的基因治疗产品外,没有其他治疗方法。在学术界研发并获批后,华尔街分析师预测Luxturna的销售额到2022年将达到3.46亿美元。尽管实际销售额低于这一初步预测(每年在1000万至5000万美元之间),但Luxturna的销售额仍超过了Roctavian。一种解释是,由于治疗双眼的费用为85万美元,Luxturna是市场上价格最低的AAV基因药物。这种中间情况很可能代表了罕见病AAV药物的商业表现,除非它确实是一个未被满足的医疗需求。
AAV未来临床成功的展望:从实验室到临床应用
利用AAV进行罕见疾病和原发性疾病的基因治疗仍存在许多挑战,提高这些产品的安全性和有效性是当务之急。首个使用AAV2衣壳进行FIX递送的静脉注射载体试验表明,肝脏酶水平出现无症状升高,随后循环中的FIX水平降低;基于这一结果,研究人员推测,对衣壳的免疫反应是表达量下降的原因。后来,Nathwani的试验表明,使用类固醇干预可阻止肝酶水平升高,并保持FIX表达。为了尽量减少达到有效剂量所需的剂量,科学家们致力于优化载体以提高表达。对于分泌蛋白而言,这似乎是一种可行的方法。scAAV载体的开发为适合这些分子独特载体(SMA)包装的转基因提供了可行的答案。然而Kattenhorn等人提出了一个需谨慎对待的关于AAV基因疗法的警示:这些作者认为,增强表达,尤其是通过密码子优化来实现的增强表达,可能会导致不良后果。这一观点有时会被忽视,但事实已经明确:一些罕见密码子存在于对蛋白质正确折叠至关重要的区域中,而这些罕见密码子能够促进蛋白质的共翻译折叠。共翻译转运穿过细胞膜(例如分泌蛋白)可能需要这些罕见密码子的存在来减缓翻译,从而激活信号识别颗粒与前导肽结合并将其与膜关联。将罕见密码子替换为常见的密码子可能会导致蛋白质折叠不当,从而导致蛋白质降解或聚集(包涵体),或是导致结构和翻译后修饰的较轻微变化,从而影响蛋白质的功能。在MDR1基因中发生的沉默性替换会导致对化疗药物和心血管药物的敏感性降低,而在COMP基因中发生的沉默性替换会改变对疼痛的敏感性,这在Kimchi-Sarfaty等人及Nackley等人的研究中已有阐述。《科学美国人(Scientific American)》早在2009年就估计,至少有50种沉默性突变通过各种机制与人类疾病有关。在所有这些机制中,细胞内错误折叠蛋白质的积累可能导致未折叠蛋白反应,并产生严重的毒性。
AAV基因疗法试验中的致命毒性较为罕见,似乎与高剂量的静脉注射治疗产品有关。Duan已对迄今为止所有已知由基因疗法载体引起的致命毒性病例进行了仔细审查,发现了与基因疗法载体相关的肝衰竭、肾衰竭、心衰以及最近的肺衰竭等病例。在最新的案例中,有观点提出,细胞因子诱导的毛细血管渗漏综合征可能是注射了CRD-TMH-001载体后出现的肺功能衰竭的原因;该载体携带基于CRISPR的转录激活因子。肝毒性与8例死亡病例有关:4例患有杜氏肌营养不良症(DMD)的患者接受了剂量为1.1e+14 vg/kg的Zolgensma治疗,另外4例患有X连锁性肌管性肌病(XLMTM)的患者接受了剂量为5e13-3e14 vg/kg的AT132治疗。衣壳驱动的免疫反应可能是Zolgensma案例中的原因,而先前存在的肝胆疾病则与XLMTM案例有关。另外一起在使用Zolgensma治疗后死亡的病例与涉及补体激活的TMA有关,并伴有由于葡萄球菌感染引起的败血症。由先天免疫反应导致的心脏病变是接受PF-06939926(剂量为2e14 vg/kg)治疗的一名DMD患者的死因。肝毒性在20%至90%的患者中均有出现,从低剂量下无症状患者(AAV5,AAV6,AAV-LK03,SPARK100,4.5e11-6e13 vg/kg)到对类固醇有反应的肝功能衰竭患者(AAV9,1.1e14 vg/kg)再到肝功能衰竭并死亡的患者(AAV8,1.3e14-3.5e14 vg/kg);其严重程度与载体剂量成正比。然而,作者忽略了一个关于无症状和更严重肝功能结果之间显著差异的问题,这一差异对于解读可能非常重要:所有无症状患者均为成年人,而那些出现更严重结果的患者均为儿童患者。特发性门脉高压症较为罕见,仅在剂量为1.1e14-3e14 vg/kg且使用AAV9衣壳(无论转录因子为何,包括SMA、两种不同的DMD、Danon)的情况下观察到,所有病例均通过血液透析和依库珠单抗进行治疗。有一例特发性门脉高压症则是在一个剂量下观察到的。
结论
尽管我们对于影响基因成功传递的众多变量的理解仍处于初期阶段,但社会和经济因素对于该领域的成功同样重要,其重要性不亚于最优质的衣壳载体或最新能解决技术难题的启动子元件。作为致力于实现从实验室到临床应用这一承诺的研究人员,我们已经取得了极高的成就。例如,脊髓性肌萎缩症于1891年被发现,导致脊髓性肌萎缩症的SMN1基因于1995年被发现;最近一名新生儿在出生第4天就被诊断出患有脊髓性肌萎缩症,并在第7天接受了FDA批准的药物治疗。随着超过300项临床试验的进行,交流临床经验(例如,处理肝脏毒性等问题)现在已成为实现治疗成功的关键驱动力。
参考文献
Suarez-Amaran L, Song L, Tretiakova AP, Mikhail SA, Samulski RJ.AAV vector development, back to the future. Mol Ther.2025May7;33(5):1903-1936. doi:10.1016/j.ymthe.2025.03.064. Epub2025Apr3. PMID:40186350; PMCID: PMC12126818.

地址:中国武汉东湖高新区光谷七路128号 市场:17720522078 人事行政:027-62439686 邮箱:marketing@genevoyager.com
BD 商务总台:17720514121 /13886000399(BD 经理) /17720515371(BD 经理)
本公司所有产品仅供实验科研使用,不用于人体疾病治疗及临床诊断。
地址:中国武汉东湖高新区光谷七路128号 市场:17720522078 人事行政:027-62439686 邮箱:marketing@genevoyager.com
BD 商务总台:17720514121 /13886000399(BD 经理) /17720515371(BD 经理)
本公司所有产品仅供实验科研使用,不用于人体疾病治疗及临床诊断。
